


Age‐induced hair greying – the multiple effects of oxidative stress
An obvious sign of ageing is hair greying, or the loss of pigment production and deposition within the hair shafts. Numerous mechanisms, acting at different levels and follicular locations, contribute to hair greying, ranging from melanocyte stem cells defects to follicular melanocyte death. One key issue that is in common to these processes is oxidative damage. At the hair follicle stem cells niche, oxidative stress, accelerated by B‐cell lymphoma 2 gene (BCL‐2) depletion, leads to selective apoptosis and diminution of melanocyte stem cells, reducing the repopulation of newly formed anagen follicles. Melanotic bulbar melanocytes express high levels of BCL‐2 to enable survival from melanogenesis‐ and ultraviolet A (UVA)‐induced reactive oxygen species (ROS) attacks. With ageing, the bulbar melanocyte expression of anti‐oxidant proteins such as BCL‐2, and possibly TRP‐2, is reduced, and the dedicated enzymatic anti‐oxidant defence system throughout the follicle weakens, resulting in enhanced oxidative stress. A marked reduction in catalase expression and activity results in millimolar accumulation of hydrogen peroxide, contributing to bulbar melanocyte malfunction and death. Interestingly, amelanotic melanocytes at the outer root sheath (ORS) are somewhat less affected by these processes and survive for longer time even within the white, ageing hair follicles. Better understanding of the overtime susceptibility of melanocytes to oxidative stress at the different follicular locations might yield clues to possible therapies for the prevention and reversal of hair greying.
Oxidative Stress in Ageing of Hair
Experimental evidence supports the hypothesis that oxidative stress plays a major role in the ageing process. Reactive oxygen species are generated by a multitude of endogenous and environmental challenges. Reactive oxygen species or free radicals are highly reactive molecules that can directly damage cellular structural membranes, lipids, proteins, and DNA. The body possesses endogenous defence mechanisms, such as antioxidative enzymes and non-enzymatic antioxidative molecules, protecting it from free radicals by reducing and neutralizing them. With age, the production of free radicals increases, while the endogenous defence mechanisms decrease. This imbalance leads to the progressive damage of cellular structures, presumably resulting in the ageing phenotype. Ageing of hair manifests as decrease of melanocyte function or graying, and decrease in hair production or alopecia. There is circumstantial evidence that oxidative stress may be a pivotal mechanism contributing to hair graying and hair loss. New insights into the role and prevention of oxidative stress could open new strategies for intervention and reversal of the hair graying process and age-dependent alopecia.
Age-induced hair graying (canities), or the age-induced loss of melanin synthesis and deposition within the hair shafts, is a noticeable and undesired sign of the aging process. Numerous mechanisms contribute to age-induced hair graying, affecting both follicular and stem cell melanocytes and acting at different follicular locations. Many of these processes are induced, directly or indirectly, by oxidative insults and damage. Melanin-producing bulbar melanocytes express high levels of BCL-2 to survive reactive oxygen species (ROS) attacks, which are induced by the melanogenic process itself and by ultraviolet A (UVA) irradiation. With aging, the expression of BCL-2, and possibly of TRP-2, is reduced, and the endogenous, enzymatic antioxidant defense system declines, resulting in greater oxidative stress. In particular, catalase expression and activity are markedly reduced with aging, leading to millimolar accumulation of hydrogen peroxide within the hair follicle and contributing to bulbar melanocyte failure and death. Additionally, exposure of melanocyte stem cells to cumulative oxidative damage, combined with reduced BCL-2 protective levels, results in apoptosis and therefore decreases the number of melanocytes that could repopulate the newly formed anagen follicles. Altogether, oxidative stress may contribute to age-induced hair graying via multiple pathways. Better understanding of the different processes, sources, and types of oxidative stress within the follicular environment, and the different susceptibilities of melanocytes to oxidative stress at the different follicular locations, might yield clues to possible interventions for prevention or reversal of hair graying.
Here we provide unique evidence for oxidative stress induced loss of melanocytes from the human hair follicle during aging. In detail, we show for the first time that:1)a decreased number of viable melanocytes in the aging hair follicle bulge and bulb and an increased incidence of hair bulb melanocyte apoptosis in aging individuals are associated with oxidative stress in the pigmentary unit;2)the aging hair follicle is characterized by the absence of oxidative stress-protectors, such as Bcl-2, and melanocyte growth factors, such as c-Kit;3)a higher frequency of oxidative stress associated mitochondrial DNA damage occurs in graying hair follicles, while unpigmented hair follicles prove to be not “older” than pigmented hair follicles;4)melanocytes of the pigmentary unit are highly and selectively susceptible to exogenous oxidative stress damage.One major route, by which oxidative stress leads to permanent melanocyte damage, appears to pass by the mitochondria, since their DNA is not so well protected as genomic DNA. The accumulation of mutations, therefore, correlates with age and is indicative of general exposure and generation of oxidative stress, (39⤻, 40)⤻caused for example by psychoemotional stress, inflammation, UV-light, and others. In summary, our findings support the proposed hypothesis of a “free radical theory of graying” and suggest that melanocytes in the hair follicle are highly susceptible to endogenous oxidative stress. Exogenous oxidative stress can trigger and hasten this process and provoke permanent damage selectively and prematurely in hair bulb melanocytes.
Age-related hair pigment loss.
Humans are social animals that communicate disproportionately via potent genetic signals imbued in the skin and hair, including racial, ethnic, health, gender, and age status. For the vast majority of us, age-related hair pigment loss becomes the inescapable signal of our disappearing youth. The hair follicle (HF) pigmentary unit is a wonderful tissue for studying mechanisms generally regulating aging, often before this becomes evident elsewhere in the body. Given that follicular melanocytes (unlike those in the epidermis) are regulated by the hair growth cycle, this cycle is likely to impact the process of aging in the HF pigmentary unit. The formal identification of melanocyte stem cells in the mouse skin has spurred a flurry of reports on the potential involvement of melanocyte stem cell depletion in hair graying (i.e., canities). Caution is recommended, however, against simple extrapolation of murine data to humans. Regardless, hair graying in both species is likely to involve an age-related imbalance in the tissue’s oxidative stress handling that will impact not only melanogenesis but also melanocyte stem cell and melanocyte homeostasis and survival. There is some emerging evidence that the HF pigmentary unit may have regenerative potential, even after it has begun to produce white hair fibers. It may therefore be feasible to develop strategies to modulate some aging-associated changes to maintain melanin production for longer.
Chronic inflammation induces telomere dysfunction and accelerates ageing
Chronic inflammation is associated with normal and pathological ageing. Here we show that chronic, progressive low-grade inflammation induced by knockout of the nfkb1subunit of the transcription factor NF-κB induces premature ageing in mice. We also show that these mice have reduced regeneration in liver and gut. nfkb1−/−fibroblasts exhibit aggravated cell senescence because of an enhanced autocrine and paracrine feedback through NF-κB, COX-2 and ROS, which stabilizes DNA damage. Preferential accumulation of telomere-dysfunctional senescent cells in nfkb1−/−tissues is blocked by anti-inflammatory or antioxidant treatment of mice, and this rescues tissue regenerative potential. Frequencies of senescent cells in liver and intestinal crypts quantitatively predict mean and maximum lifespan in both short- and long-lived mice cohorts. These data indicate that systemic chronic inflammation can accelerate ageing via ROS-mediated exacerbation of telomere dysfunction and cell senescence in the absence of any other genetic or environmental factor.
Aging is associated with circulating cytokine dysregulation
Aging has a significant impact on the production of circulating cytokines in healthy individuals. The circulating cytokine milieu may contribute to the development of age-restricted conditions. ► Aging has a significant impact on the production of circulating cytokines. ► Cytokines dysregulation is demonstrated ex vivo but not after in vitro activation. ► Pro- and anti-inflammatory cytokines correlate with aging. ► Th1 cytokines increase whereas Th17 cytokines decrease with age.
Repigmentation and new growth of hairs after anti–interleukin-17 therapy
Repigmentation of hairs is a rare event that has been reported after inflammatory processes, exposure to X-irradiation and psoralen and ultraviolet A, electron beam therapy, and the intake of some drugs.1 We report on a patient with psoriasis who experienced darkening and noticeable increase in scalp hair while he was receiving anti–interleukin (IL)-17 therapy.
Using RNA in situ hybridization analysis, we have characterized the expression domains of the four known members of the FGF receptor-tyrosine kinase gene family in the murine hair follicle at various stages of the hair growth cycle. During anagen, we detected Fgfr1 RNA in the dermal papilla, Fgfr2 RNA in hair matrix cells near the dermal papilla, Fgfr3 RNA in pre-cuticle cells in the periphery of the hair bulb, and Fgfr4 RNA in cells in the periphery of the hair bulb and also in the inner and outer root sheath in the lower half of the follicle neck. No RNA expression of these genes was detected during late catagen or telogen. We have previously shown that Fgf5 is expressed in the outer root sheath in the transient portion of the follicle (Hébert et al. [1994] Cell 78:1017-1025). In the present study we have also assayed for the expression of six other members of the FGF ligand gene family, Fgf3, Fgf4, Fgf6, Fgf7, Fgf8, and Fgf9. Among these FGF genes, only Fgf7 was found to be expressed in the hair follicle. Fgf7 RNA is localized to the dermal papilla during anagen, but expression is down-regulated by the late-anagen VI stage. We have also demonstrated that addition of FGF5 protein to the culture medium changes the behavior of dermal papilla cells in vitro, indicating that they are capable of responding to FGF5. Together with previously published data, these results provide a complete analysis of FGF ligand and FGF receptor-tyrosine kinase gene expression in the hair follicle, and suggest that FGF signalling may have several functions in the hair growth cycle
Hair shaft producing cells were proliferated only in the anagen. Many mediators were involved in starting, sustaining and terminating anagen. Among them, we focused on the growth factor FGF-7 and FGF-5. Our results reveal the hair growth enhancing effects of EP were related with the anagen regulating growth factors. mTOR activating the potential of EP in HDPs was signifying that it exerts the positive role in the proliferation of follicular cells during anagen.
FGF5 is a crucial regulator of hair length in humans
Hair length varies dramatically on different body sites and also varies between individuals. Thus, hair length is a quantitative trait, suggesting inherited differences. In this study, we obtained DNA from families segregating excessively long eyelashes consistent with an autosomal recessive trait. We identified mutations in a single gene, fibroblast growth factor 5 (FGF5), which was homozygous in affected family members only. FGF5 has previously been implicated as a regulator of hair lengths in mammals, with mutations resulting in the well-described angoraphenotype. However, until now a human counterpart to this phenotype remained elusive. Here, we present, to our knowledge, the first human counterpart of the angoraphenotype, showing that FGF5 underlies trichomegaly and is a crucial regulator of hair growth in humans.
Widespread expression of the transcription factor, nuclear factor (erythroid‐derived 2)‐like 2 (NRF2), which maintains redox homeostasis, has recently been identified in the hair follicle (HF). Small molecule activators of NRF2 may therefore be useful in the management of HF pathologies associated with redox imbalance, ranging from HF greying and HF ageing via androgenetic alopecia and alopecia areata to chemotherapy‐induced hair loss. Indeed, NRF2 activation has been shown to prevent peroxide‐induced hair growth inhibition. Multiple parameters can increase the levels of reactive oxygen species in the HF, for example melanogenesis, depilation‐induced trauma, neurogenic and autoimmune inflammation, toxic drugs, environmental stressors such as UV irradiation, genetic defects and aging‐associated mitochondrial dysfunction. In this review, the potential mechanisms whereby NRF2 activation could prove beneficial in treatment of redox‐associated HF disorders are therefore discussed.
The impact of oxidative stress on hair
Oxidative stress reflects an imbalance between the systemic manifestation of reactive oxygen species and a biological system’s ability to detoxify the reactive intermediates or to repair the resulting damage. Reactive oxygen species or free radicals are highly reactive molecules that can directly damage lipids, proteins, and DNA. They are generated by a multitude of endogenous and environmental challenges, while the body possesses endogenous defense mechanisms. With age, production of free radicals increases, while the endogenous defense mechanisms decrease. This imbalance leads to progressive damage of cellular structures, presumably resulting in the aging phenotype. While the role of oxidative stress has been widely discussed in skin aging, little focus has been placed on its impact on hair condition. Moreover, most literature on age‐related hair changes focuses on alopecia, but it is equally important that the hair fibers that emerge from the scalp exhibit significant age‐related changes that have equal impact on the overall cosmetic properties of hair. Sources of oxidative stress with impact on the pre‐emerging fiber include: oxidative metabolism, smoking, UVR, and inflammation from microbial, pollutant, or irritant origins. Sources of oxidative stress with impact on the post‐emerging fiber include: UVR (enhanced by copper), chemical insults, and oxidized scalp lipids. The role of the dermatologist is recognition and treatment of pre‐ and post‐emerging factors for lifetime scalp and hair health.
Repigmentation of hair following adalimumab therapy
Repigmentation of canities, or age-related grey or white hair, is a rare occurrence. Generalized repigmentation of grey-white hair has been reported following inflammatory processes,[1] and heterochromia (localized patches of hair repigmentation) is even more unusual, reported in association with medication use and malignancy.Tumor necrosis factor (TNF) inhibitors are increasingly utilized medications for inflammatory disorders, including psoriasis, rheumatoid arthritis, and inflammatory bowel disease. Hair loss, or alopecia, has been described among the side effects of these medications,[2] but changes in hair pigmentation in association with this class of drugs have not previously been reported. We describe a patient with hair repigmentation associated with adalimumab therapy.
A Comment on the Science of Hair Aging
In contrast to the skin, aging of the hair has seemingly only recently found the attention of dermatological meetings, mainly promoted by the cosmetic industry for marketing purposes. In fact, basic scientists interested in the biology of hair growth and pigmentation have for some time already exposed the hair follicle as a highly accessible model with unique opportunities for the study of age-related effects. As a result, the science of hair aging focuses on two main streams of interest: the esthetic problem of aging hair and its management, in terms of age-related effects on hair color, quantity, and quality; and the biological problem of aging hair, in terms of microscopic, biochemical, and molecular changes underlying the aging process. Ultimately, the aim of hair anti-aging is to delay, lessen, or reverse the effects of aging on hair. According to the complex nature of the aging process, the treatment for lifetime scalp and hair health has to be holistic to include the multitude of contributing factors in a polyhedral and patient-specific manner. It comprises both medical treatments and hair cosmetics. Accordingly, the discovery of pharmacological targets and the development of safe and effective drugs for treatment of hair loss indicate strategies of the drug industry for maintenance of hair growth and quantity, while the hair care industry has become capable of delivering active compounds directed toward meeting the consumer demand for maintenance of hair cosmesis and quality. “Where there’s life, there’s hope” (Ecclesiastes 9:3-5).
Three Streams for the Mechanism of Hair Graying
Hair graying is an obvious sign of human aging. Although graying has been investigated extensively, the mechanism remains unclear. Here, we reviewed previous studies on the mechanism of graying and seek to offer some new insights. The traditional view is that hair graying is caused by exhaustion of the pigmentary potential of the melanocytes of hair bulbs. Melanocyte dysfunction may be attributable to the effects of toxic reactive oxygen species on melanocyte nuclei and mitochondria. A recent study suggests that bulge melanocyte stem cells (MSCs) are the key cells in play. Graying may be caused by defective MSC self-maintenance, not by any deficiency in bulbar melanocytes. Our previous study suggested that graying may be principally attributable to active hair growth. Active hair growth may produce oxidative or genotoxic stress in hair bulge. These internal stress may cause eventually depletion of MSC in the hair follicles. Taken together, hair graying may be caused by MSC depletion by genotoxic stress in the hair bulge. Hair graying may also be sometimes caused by dysfunction of the melanocytes by oxidative stress in the hair bulb. In addition, hair graying may be attributable to MSC depletion by active hair growth.
Pharmacologic interventions in aging hair
The appearance of hair plays an important role in people’s overall physical appearance and self-perception. With today’s increasing life-expectations, the desire to look youthful plays a bigger role than ever. The hair care industry has become aware of this and is delivering active products directed towards meeting this consumer demand. The discovery of pharmacological targets and the development of safe and effective drugs also indicate strategies of the drug industry for maintenance of healthy and beautiful hair. Hair aging comprises weathering of the hair shaft, decrease of melanocyte function, and decrease in hair production. The scalp is subject to intrinsic and extrinsic aging. Intrinsic factors are related to individual genetic and epigenetic mechanisms with interindividual variation: prototypes are familial premature graying, and androgenetic alopecia. Currently available pharmacologic treatment modalities with proven efficacy for treatment of androgenetic alopecia are topical minoxidil and oral finasteride. Extrinsic factors include ultraviolet radiation and air pollution. Experimental evidence supports the hypothesis that oxidative stress also plays a role in hair aging. Topical anti-aging compounds include photoprotectors and antioxidants. In the absence of another way to reverse hair graying, hair colorants remain the mainstay of recovering lost hair color. Topical liposome targeting for melanins, genes, and proteins selectively to hair follicles are currently under investigation.
Stress has long been discussed controversially as a cause of hair loss. However, solid proof of stress‐induced hair growth inhibition had long been missing. If psychoemotional stress can affect hair growth, this must be mediated via definable neurorendocrine and/or neuroimmunological signaling pathways. Revisiting and up‐dating relevant background data on neural mechanisms of hair growth control, we sketch essentials of hair follicle (HF) neurobiology and discuss the modulation of murine hair growth by neuropeptides, neurotransmitters, neurotrophins, and mast cells. Exploiting an established mouse model for stress, we summarize recent evidence that sonic stress triggers a cascade of molecular events including plasticity of the peptidergic peri‐ and interfollicular innervation and neuroimmune crosstalk. Substance P (SP) and NGF (nerve growth factor) are recruited as key mediators of stress‐induced hair growth‐inhibitory effects. These effects include perifollicular neurogenic inflammation, HF keratinocyte apoptosis, inhibition of proliferation within the HF epithelium, and premature HF regression (catagen induction). Intriguingly, most of these effects can be abrogated by treatment of stressed mice with SP‐receptor neurokinin‐1 receptor (NK‐1) antagonists or NGF‐neutralizing antibodies – as well as, surprisingly, by topical minoxidil. Thus there is now solid in vivo‐evidence for the existence of a defined brain‐ HF axis. This axis can be utilized by psychoemotional and other stressors to prematurely terminate hair growth. Stress‐induced hair growth inhibition can therefore serve as a highly instructive model for exploring the brain‐skin connection and provides a unique experimental model for dissecting general principles of skin neuroendocrinology and neuroimmunology well beyond the HF.
The appearance of hair plays an important role in people’s overall physical appearance and self‐perception. With today’s increasing life expectation, the desire to look youthful plays a bigger role than ever. The hair care industry has become aware of this and also more capable to deliver active products that are directed toward meeting this consumer demand. The discovery of pharmacological targets and the development of safe and effective drugs also indicate strategies of the drug industry for maintenance of healthy and beautiful hair. Hair aging comprises weathering of the hair shaft and aging of the hair follicle. The latter manifests as decrease of melanocyte function or graying, and decrease in hair production in androgenetic and senescent alopecia. The scalp is also subject to intrinsic or physiologic aging and extrinsic aging caused by external factors. Intrinsic factors are related to individual genetic and epigenetic mechanisms with interindividual variation. Prototypes are familial premature graying and androgenetic alopecia. Extrinsic factors include ultraviolet radiation and smoking. Experimental evidence supports the hypothesis that oxidative stress plays a role in skin and hair aging. Topical anti‐aging compounds for hair include humefactants, hair conditioners, photoprotectors, and antioxidants. Current available treatment modalities with proven efficacy for treatment of androgenetic alopecia are topical minoxidil, oral finasteride, and autologous hair transplantation. In the absence of another way to reverse hair graying, hair colorants are the mainstays of recovering lost hair color. Topical liposome targeting for melanins, genes, and proteins selectively to hair follicles are under current investigation.
Hair cycle and hair pigmentation: dynamic interactions and changes associated with aging
The tight coupling of hair follicle melanogenesis to the hair growth cycle dramatically distinguishes follicular melanogenesis from the continuous melanogenesis of the epidermis. Cyclic re-construction of an intact hair follicle pigmentary unit occurs optimally in all scalp hair follicles during only the first 10 hair cycles, i.e. by approximately 40 years of age. Thereafter there appears to be a genetically regulated exhaustion of the pigmentary potential of each individual hair follicle leading to the formation of true gray and white hair. Pigment dilution results primarily from a reduction in tyrosinase activity within hair bulbar melanocytes. Thereafter, sub-optimal melanocyte–cortical keratinocyte interactions, and defective migration of melanocytes from a reservoir in the upper outer root sheath to the pigment-permitting microenvironment close to the follicular papilla of the hair bulb, will all disrupt normal function of the pigmentary unit. Evidence from studies on epidermal melanocyte aging suggest that reactive oxygen species-mediated damage to nuclear and mitochondrial DNA may lead to mutation accumulation in bulbar melanocytes. Parallel dysregulation of anti-oxidant mechanisms or pro/anti-apoptotic factors is also likely to occur within the cells. Pigment loss in canities may also affect keratinocyte proliferation and differentiation, providing the tantalizing suggestion that melanocytes in the hair follicle contribute far more that packages of pigment alone. Here, we review the current state of knowledge of the development, regulation and control of the aging human hair follicle pigmentary system in relation with hair cycling. The exploitation of recently available methodologies to manipulate hair follicle melanocytes in vitro, and the observations that melanocytes remain in senile white hair follicles that can be induced to pigment in culture, raises the possibility of someday reversing canities. The perspective of rejuvenation of the whole hair follicle apparatus are still part of the dream but optimising its functional properties is clinically relevant and is close to reality. Finally as hair color influences its visibility when optical methods such as scalp photography are used to count hair fibers, the attention is drawn to possible interpretations of statistically significant changes in visible hair. Such changes may not exclusively be related to improved hair growth itself but also to changes in natural hair color that makes the hair more visible with the method used to count hairs.
Human hair pigmentation – biological aspects
Skin and hair colour contribute significantly to our overall visual appearance and to social/sexual communication. Despite their shared origins in the embryologic neural crest, the hair follicle and epidermal pigmentary units occupy distinct, although open, cutaneous compartments. They can be distinguished principally on the basis of the former’s stringent coupling to the hair growth cycle compared with the latter’s continuous melanogenesis. The biosynthesis of melanin and its subsequent transfer from melanocyte to hair bulb keratinocytes depend on the availability of melanin precursors and on a raft of signal transduction pathways that are both highly complex and commonly redundant. These signalling pathways can be both dependent and independent of receptors, act through auto‐, para‐ or intracrine mechanisms and can be modified by hormonal signals. Despite many shared features, follicular melanocytes appear to be more sensitive than epidermal melanocytes to ageing influences. This can be seen most dramatically in hair greying/canities and this is likely to reflect significant differences in the epidermal and follicular microenvironments. The hair follicle pigmentary unit may also serve as an important environmental sensor, whereby hair pigment contributes to the rapid excretion of heavy metals, chemicals and toxins from the body by their selective binding to melanin; rendering the hair fibre a useful barometer of exposures. The recent availability of advanced cell culture methodologies for isolated hair follicle melanocytes and for intact anagen hair follicle organ culture should provide the research tools necessary to elucidate the regulatory mechanisms of hair follicle pigmentation. In the longer term, it may be feasible to develop hair colour modifiers of a biological nature to accompany those based on chemicals.
Histopathology of aging of the hair follicle
Hair follicles experience several changes with aging, the most noticeable of which is graying of the hair shaft due to loss of melanin. Additional changes in the diameter and length of the hair have contributed to the concept of senescent alopecia, which is different from androgenetic alopecia according to most. Graying happens in most individuals, although in different grades and starting at different ages. It is related to a decrease in the number and activity of the melanocytes of the hair bulb, which eventually completely disappear from the bulb of the white hair. Residual non‐active melanocytes remain in the outer root sheath and in the bulge, which allows for repigmentation of the hair under certain stimuli or conditions.
A review of the etiologies, clinical characteristics, and treatment of grey hair
Hair pigmentation is regulated by follicular melanogenesis, in which the process consists of melanin formation and transfer to keratinocytes in the hair shaft. Human hair follicles contain two types of melanin: the brown‐black eumelanin and yellow‐red pheomelanin. Eumelanin is commonly present in black and brown hair while pheomelanin is found in auburn and blonde hair. Hair follicle melanogenesis is under cyclical control and is concurrently coupled to hair growth. Many factors including intrinsic and extrinsic factors affect the follicular melanogenesis. Though many studies have been conducted to identify the pathogenesis and regulation of hair pigmentation, the etiology of canities and hair pigmentation is still unclear. The pathogenesis of canities or gray hair is believed to occur either from insufficient melanin formation due to melanocyte degeneration or a defect in melanosomal transfer. Canities is an aging sign which often interferes with one’s socio‐cultural adjustment. On the other hand, premature canities correlate with diseases such as osteopenia and cardiovascular disease. Risk factors associated with canities are not only genetic but also external causes. For example, smoking, alcohol consumption, and stress are among the most common factors. Camouflage techniques are still used as the primary treatment of canities. Further treatments for canities are being developed to achieve the true reversal of hair pigmentation.
Aging of the Hair Follicle Pigmentation System
Skin and hair phenotypes are powerful cues in human communication. They impart much information, not least about our racial, ethnic, health, gender and age status. In the case of the latter parameter, we experience significant change in pigmentation in our journey from birth to puberty and through to young adulthood, middle age and beyond. The hair follicle pigmentary unit is perhaps one of our most visible, accessible and potent aging sensors, with marked dilution of pigment intensity occurring long before even subtle changes are seen in the epidermis. This dichotomy is of interest as both skin compartments contain melanocyte subpopulations of similar embryologic (i.e., neural crest) origin. Research groups are actively pursuing the study of the differential aging of melanocytes in the hair bulb versus the epidermis and in particular are examining whether this is in part linked to the stringent coupling of follicular melanocytes to the hair growth cycle. Whether some follicular melanocyte subpopulations are affected, like epidermal melanocytes, by UV irradiation is not yet clear. A particular target of research into hair graying or canities is the nature of the melanocyte stem compartment and whether this is depleted due to reactive oxygen species-associated damage, coupled with an impaired antioxidant status, and a failure of melanocyte stem cell renewal. Over the last few years, we and others have developed advanced in vitro models and assay systems for isolated hair follicle melanocytes and for intact anagen hair follicle organ culture which may provide research tools to elucidate the regulatory mechanisms of hair follicle pigmentation. Long term, it may be feasible to develop strategies to modulate some of these aging-associated changes in the hair follicle that impinge particularly on the melanocyte populations.
Autocrine and paracrine factors are produced by balding dermal papilla (DP) cells following dihydrotestosterone (DHT)-driven alterations and are believed to be key factors involved in male pattern baldness. Herein we report that the IL-6 is upregulated in balding DP cells compared with non-balding DP cells. IL-6 was upregulated 3 hours after 10-100 nM DHT treatment, and ELISA showed that IL-6 was secreted from balding DP cells in response to DHT. IL-6 receptor (IL-6R) and glycoprotein 130 (gp130) were expressed in follicular keratinocytes, including matrix cells. Recombinant human IL-6 (rhIL-6) inhibited hair shaft elongation and suppressed proliferation of matrix cells in cultured human hair follicles. Moreover, rhIL-6 injection into the hypodermis of mice during anagen caused premature onset of catagen. Taken together, our data strongly suggest that DHT-inducible IL-6 inhibits hair growth as a paracrine mediator from the DP.
Cytokines and Other Mediators in Alopecia Areata
Alopecia areata, a disease of the hair follicles with multifactorial etiology and a strong component of autoimmune origin, has been extensively studied as far as the role of several cytokines is concerned. So far, IFN-, interleukins, TNF-, are cytokines that are well known to play a major role in the pathogenesis of the disease, while several studies have shown that many more pathways exist. Among them, MIG, IP-10, BAFF, HLA antigens, MIG, as well as stress hormones are implicated in disease onset and activity. Within the scope of this paper, the authors attempt to shed light upon the complexity of alopecia areata underlying mechanisms and indicate pathways that may suggest future treatments.
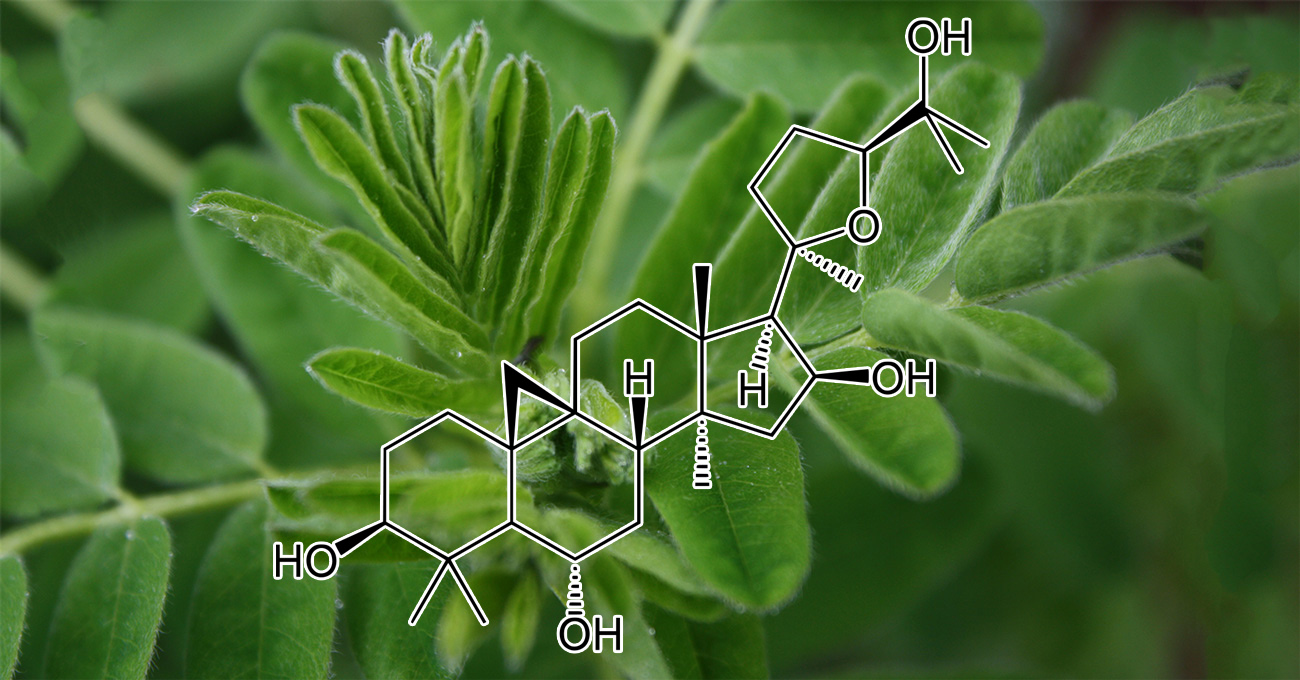
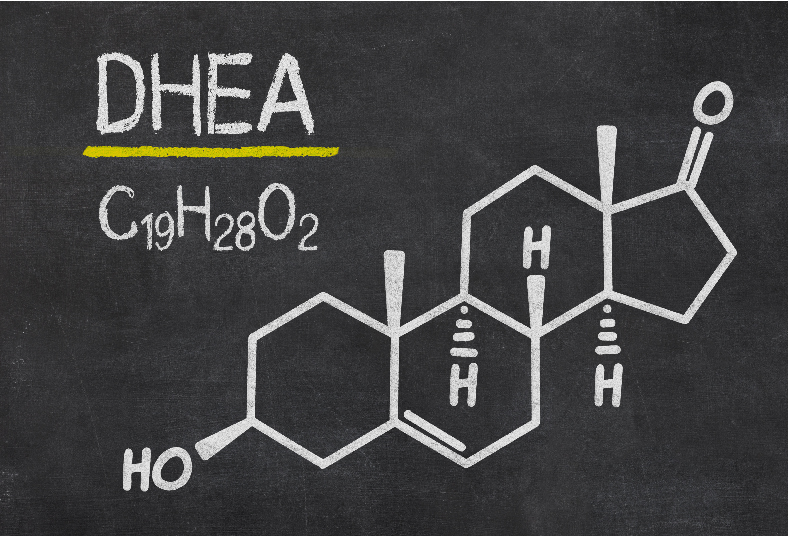
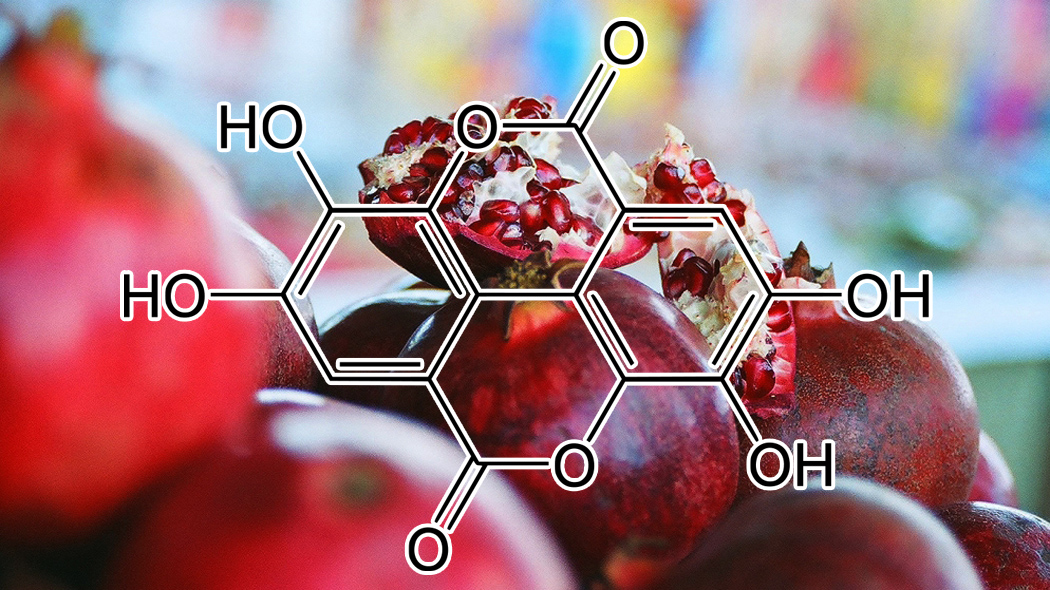
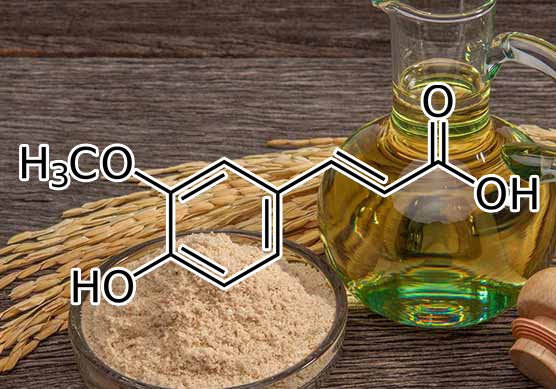
INGREDIENTS:






















































Reviews
There are no reviews yet.